
【ACS Catal.】5α还原酶的快速运动及其对酶动力学的影响

摘要
酶催化效率的背后,是残基之间精妙调控的动态相互作用,它们协同配合共同推进反应。甾体5α-还原酶2型(SRD5A2)催化NADPH依赖的睾酮还原为二氢睾酮,该反应通过连续的氢化物与质子转移实现。本研究填补了理解该催化机制方面的基本空白——活化能垒、速率促进动力学及静电贡献——这些迄今均未被表征。通过采用过渡路径采样的QM/MM模拟,我们揭示了分子运动如何影响催化反应的动力学。我们发现,Tyr33与L224两个残基对供体具有压缩效应,这不仅显著改变了自由能垒,还能够扰动局部电场,从而支持了预组织理论。在此基础上,通过广泛的通量分析(committor analysis),我们进一步鉴定出6个额外残基——Trp53、Arg94、Cys119、Glu197、Phe223和Arg227——它们构成了一个扩展的反应坐标网络,通过偶联的静电与结构相互作用来稳定过渡态。对疾病相关突变体L224P的分析表明,L224压缩效应的丧失导致电场增强作用消失,并使能垒升高3.3−3.4 kcal/mol,从而确立了高效催化需要动力学与静电学在扩展的蛋白质结构域中实现时间上的协同配合。
1.研究背景
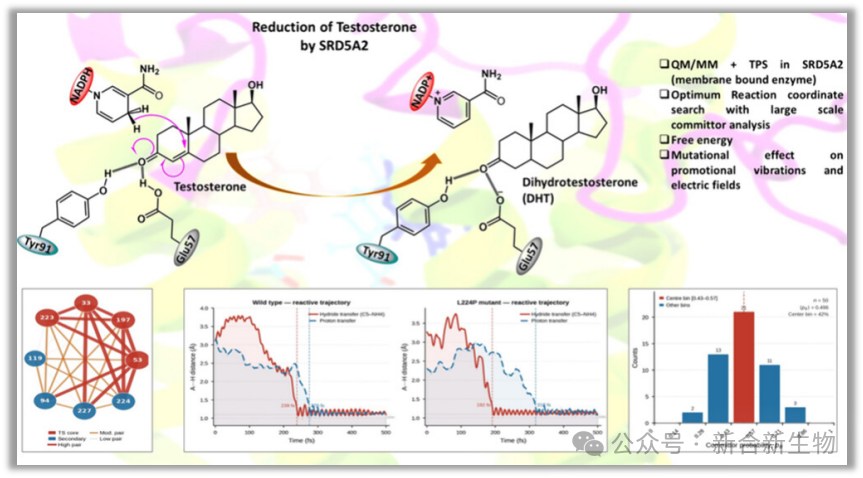
类固醇5α还原酶2型(SRD5A2)是一种由254个氨基酸组成、分子量约29.5 kDa的整合膜蛋白,由人类2p23染色体上的基因编码。该酶高度表达于内质网,含有七个跨膜α-螺旋,形成容纳底物的大内腔。SRD5A2催化NADPH依赖的睾酮还原为5α-二氢睾酮(DHT)——DHT是活性最强的天然雄激素,对雄激素受体的亲和力约为睾酮的五倍。在人体中,该酶在前列腺、精囊和生殖器皮肤等雄激素敏感组织中表达,调控局部DHT的产生,对男性性分化、前列腺发育和第二性征至关重要。SRD5A2也是经临床验证的治疗靶点——非那雄胺和度他雄胺等竞争性抑制剂通过降低前列腺内DHT水平高达90%,广泛用于良性前列腺增生和雄激素性脱发的治疗。该酶的失功能突变可导致类固醇5α还原酶2型缺乏症,目前已鉴定出114种以上不同突变,约46种功能表征变体对催化效率(Vmax/Km)有显著影响。2.研究目的
本研究旨在通过量子力学/分子力学(QM/MM)模拟结合过渡路径采样(TPS)方法,深入揭示SRD5A2催化反应中蛋白质快速运动对酶动力学的调控机制。具体目标包括:
1)阐明SRD5A2催化睾酮还原为DHT的两步反应机理中,蛋白质构象动力学如何影响化学键的断裂与形成
2)鉴定对过渡态稳定化起关键作用的残基网络
3)探究在脂质双分子层膜环境下,促速振动(Rate-Promoting Vibrations,RPVs)是否仍具有催化功能
4)通过L224P致病突变体验证动力学-静电耦合机制
3.理论基础
本研究的理论框架建立在以下核心概念之上:
1)过渡态稳定化理论:Pauling经典理论认为酶通过优先稳定过渡态而非基态来实现催化加速。
2)蛋白质动力学范式:酶并非刚性的结构支架,而是内在动态的实体,其构象波动在飞秒到毫秒时间尺度上均可直接影响反应动力学。
3)促速振动(RPVs):飞秒至皮秒时间尺度上的蛋白质快速运动,通过压缩反应中心、增强静电稳定化或预组织催化残基至最优取向等方式,系统性驱动酶-底物复合物向过渡态构型演化。
4)静电预组织机制:酶活性位点的静电环境是降低反应能垒的关键因素,动态电场调控为过渡态电荷分离提供稳定化作用。
与胞质酶(如乳酸脱氢酶LDH、嘌呤核苷磷酸化酶PNP)相比,膜结合酶的快速运动是否受到脂质环境的阻尼效应,是本研究的重要理论关切。
4.研究方法
1)体系构建:初始结构取自RCSB蛋白质数据库(PDB ID:7BW1),将晶体结构中的非那雄胺替换为睾酮。体系分为QM区和MM区:QM区包含睾酮、Tyr91、质子化的Glu57以及NADH的烟酰胺环(共100个原子),采用PM3半经验方法处理;其余部分使用CHARMM36力场描述。蛋白质-配体系统嵌入POPC脂质双分子层,在73×73×105ų的TIP3P水盒子中溶剂化,添加0.15 M KCl模拟生理盐浓度。经3000步最速下降法和2000步Newton-Raphson法能量最小化后,逐步升温至300 K,在NPT系综中平衡40 ns。
2)过渡路径采样(TPS):首先构建偏置反应轨迹——对NADPH的NH4原子施加55 kcal·mol⁻¹·Å⁻²的谐波偏置力驱动氢负离子转移,对Glu57的HE2原子施加35 kcal·mol⁻¹·Å⁻²的偏置力驱动质子转移,对Tyr91与睾酮之间施加25 kcal·mol⁻¹·Å⁻²的约束维持氢键。偏置轨迹以1 fs时间步长运行500 fs,随后用于Metropolis Monte Carlo随机游走生成了280条无偏反应轨迹。
3)自由能计算:采用反演Boltzmann方法计算氢负离子转移和质子转移两个步骤的自由能剖面。使用自举法(500次迭代)评估统计不确定性。
4)电场计算:通过叠加QM区域全部原子和底物12Å截断半径内经典蛋白质原子的静电贡献,沿睾酮C=O羰基偶极方向投影计算电场。
5.分析方法
1)Commitor分析:对候选残基进行系统性组合筛选,评估各四残基组合定义过渡态系综的质量。从8个候选残基(Tyr33、Trp53、Arg94、Cys119、Glu197、Phe223、Leu224、Arg227)中生成全部70种四残基组合,以中心区间比例和平均commitor值偏离0.5的程度为指标进行评分。对28个残基对计算加权共现矩阵和配对重要性评分,揭示残基间协同耦合关系。
2)粒子重要性评分体系:通过四维指标(TS-good率、平均中心区间比例、相对提升度、Mann-Whitney U检验)归一化组合为复合评分,建立三层级重要性分类体系。
3)角度分布分析:定义Glu57羧酸根氧原子向量与Trp53吲哚环长轴向量之间的夹角,定量分析突变对活性位点几何构型的影响。
6.研究结论
1)两步反应机制:SRD5A2催化反应分两步进行——第一步NADPH的氢负离子转移至睾酮C5(自由能能垒7.8±0.24 kcal/mol),第二步Glu57作为通用酸提供质子至C4(自由能能垒5.4±0.12 kcal/mol),生成立体特异性的5α-还原类固醇。
2)序贯压缩动力学:野生型酶中存在两次显著的残基压缩事件——Tyr33在~55 fs时压缩NADPH烟酰胺环,Leu224在80~120 fs时进一步压缩供体-受体距离从4.5Å降至~3.0Å,为氢负离子转移创造有利几何条件。
3)电场动态增强:压缩运动同时放大活性位点电场——基线~-175 MV/cm,Tyr33压缩后升至-209 MV/cm,Leu224最大压缩时达-221 MV/cm。~50 MV/cm的电场增强对应2~4 kcal/mol的能垒降低。
4)L224P突变效应:突变导致自由能能垒升高3.3±0.4 kcal/mol(氢负离子转移)和3.4±0.3 kcal/mol(质子转移),两步反应间隔时间从37 fs延长至126 fs。电场剖面平坦化,失去动态增强能力。Arg227重定向与NADPH磷酸骨架结合,Tyr33-Arg227氢键断裂,Tyr33位移(CZ-NC4距离从3.24Å增至5.5Å),Trp53从封闭构象(~10°夹角)转换为开放构象(~108°夹角)。
5)过渡态残基网络:最优四残基组合为Leu224-Tyr33-Phe223-Glu197,其中Glu197-Phe223为最重要的残基对(评分0.96)。L224P突变体中过渡态稳定网络重构为Cys119-Phe223-Arg94-Glu197组合。
7.研究创新点
1)首次实现膜结合酶SRD5A2的QM/MM-TPS模拟:采用QM/MM方法结合过渡路径采样,首次直接观测该膜蛋白活性位点化学键断裂与形成的实时动态,超越经典力场方法的局限。
2)建立"残基网络→压缩动力学→电场增强→能垒降低"的完整因果链:将压缩动力学观察、commitor分析的残基网络鉴定和时变电场计算三个维度的结果统一为单一整合催化机制。
3)发现膜环境中促速振动仍具有催化功能:尽管由于脂质双分子层的阻尼效应,SRD5A2的压缩幅度(~1.5Å)小于胞质LDH(~2.5Å),但RPVs仍然是驱动膜结合酶跨越催化能垒的重要驱动力。
4)通过致病突变体L224P实现计算预测的实验验证:L224P是已知的致病突变,本研究从分子层面揭示了该单点突变如何导致过渡态稳定网络的彻底重布线,验证了动态-静电耦合机制。
5)系统性残基网络分析框架:开发了基于comittor分析的四残基组合筛选和配对重要性评分方法,可推广至其他酶体系的反应坐标鉴定研究。
原文参考:Roy,R.K.;Antoniou,D.;Schwartz,S.D.Fast Motions in 5 Alpha Reductase and Its Impact on Enzyme Kinetics.ACS Catalysis 2026.DOI:10.1021/acscatal.6c02144.

